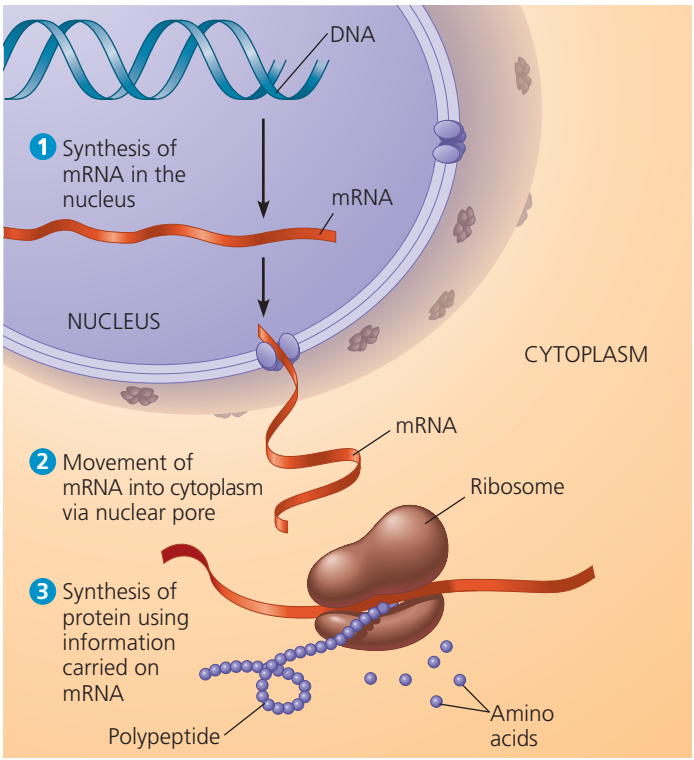
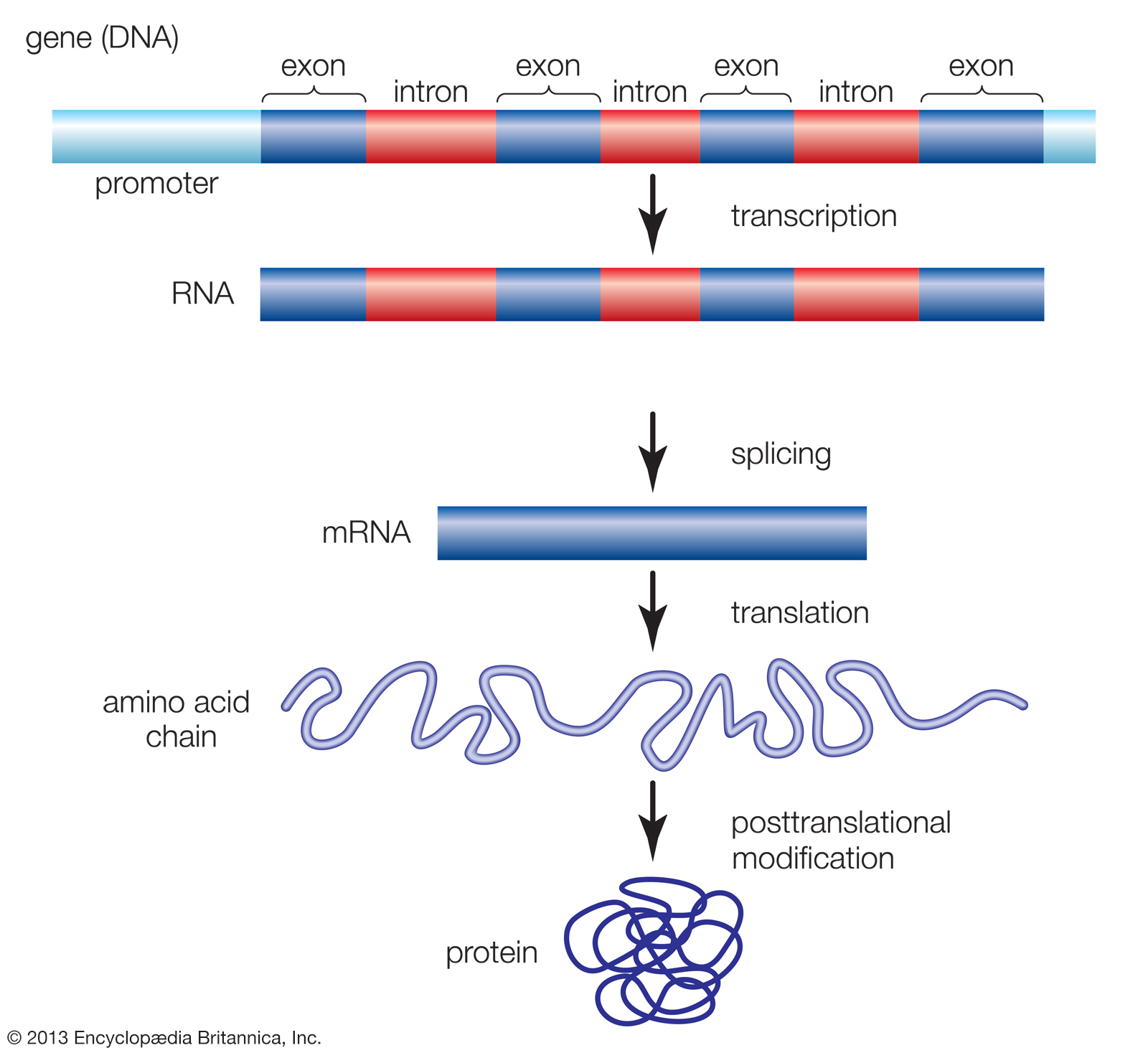
Genetik bilgi DNAdan RNAya ve ondan proteine akar.
Eşyazım süreci DNAdan RNA polimeraz aracılığıyla iletmen RNA (mRNA) zincirinin yapımıdır.
Çeviri süreci RNAdan protein üretimidir.
Ribozomal RNAnın eşyazımının büyümekte olan birincil eşyazımları gösteren elektron mikrografı. DNA telleri görüntülenemecek denli küçük yüzlerce RNA polimeraz molekülü tarafından kuşatılıdır. Her bir RNA polimeraz DNAdan dallanıyor gibi görünen bir RNA telinin eşyazımını yapmaktatır.“Begin” DNAnın yeni RNA bireşiminin başladığı noktası olan kodlayıcı telinin 5' ucunu belirtir; “end” birincil eşyazımın hemen hemen tamamlandığı 3' ucunu belirtir.
Ribozomal RNA tellerinin oluşumu ana DNA telinden dallar olarak görünürdedir. (W)
Gen anlatımı: DNA → RNA → protein. Bir ökaryotik hücrede, çekirdekteki DNA iletmen RNAnın (mRNA) bireşimini dikte ederek sitoplazmada protein üretimin programlar. RNA polimeraz kısa bir DNA kesiminin tellerini ayırır ve geçici olarak ayrılan iki DNA telinden birini kalıp olarak kullanarak tümleyici RNA eşlemini üretir.
Gen anlatımının son ürünü RNA ya da protein olabilir.
Eşyazım DNAdaki bilginin kullanımı yoluyla RNA bireşimidir.
Eşyazım sürecinde küçük bir DNA kalıbındaki kodlar yeni bir RNA molekülü olarak yeniden yazılır ya da eşlenir (eşyazım).
DNAnın protein kodlama geni üretilen RNA molekülünde tümleyici bir yolda yinelenir.
DNAdan hücrenin protein üretme düzeneğine genetik bir ileti taşıyan bu RNA molekülüne iletmen RNA (mRNA) denir.
“Eşyazım” mRNAdan başka RNA tiplerinin bireşimi için de kullanılan bir terimdir.
Eşyazım iletmen RNA (mRNA), ribozomal RNA (rRNA), aktarım RNA (tRNA) ve ek olarak özel yapısal, katalitik ve düzenleyici işlevler yerine getiren ve çeviri yapmayan (kodlamayan/noncoding) küçük RNA molekülleri (ncRNA) üretir.
Eşyazım sürecinde üretilen birçok RNA eşyazımı çeşitli değişkilere uğrar: Uç eklemeleri, baz değişkileri, kırpılma ve iç dilimlerin çıkarılması gibi işlemler işlevsiz birincil eşyazımı işlevsel moleküle dönüştürür.
RNA eşyazımı.
DNA gibi, RNA birbirine fosfodiester bağları ile bağlı dört ayrı nükleotid altbirim tipinden yapılan doğrusal bir polimerdir.
RNAdaki nükleotidler ribonükleotidlerdir (DNA nükleotidlerinde kapsanan deoksiriboz yerine RNA nükleotidleri riboz kapsar; ve bu ayrıma göre DeoksiriboNükleik Asit ve RiboNükleik Asit adlandırmaları kullanılır).
DNA A, G, C ve T bazlarını kapsarken, RNAda T yerine U bulunur.
DNA her zaman çift-telli sarmal yapıda iken, RNA tek-tellidir ve bu nedenle değişik şekillere katlanabilir, yapısal ve katalitik özellikler gösterebilir.
Polipeptidlerin birincil yapısı bir proteinin şeklini belirler.
Birincil yapı bir polipeptidin amino asit dizisidir ve gen olarak adlandırılan bir kalıtım birimi tarafından programlanır.
🛑 EŞYAZIM: DNAdan RNAya
Eşyazım RNAnın DNA-yönetimli bireşimidir: Yakından bir bakış
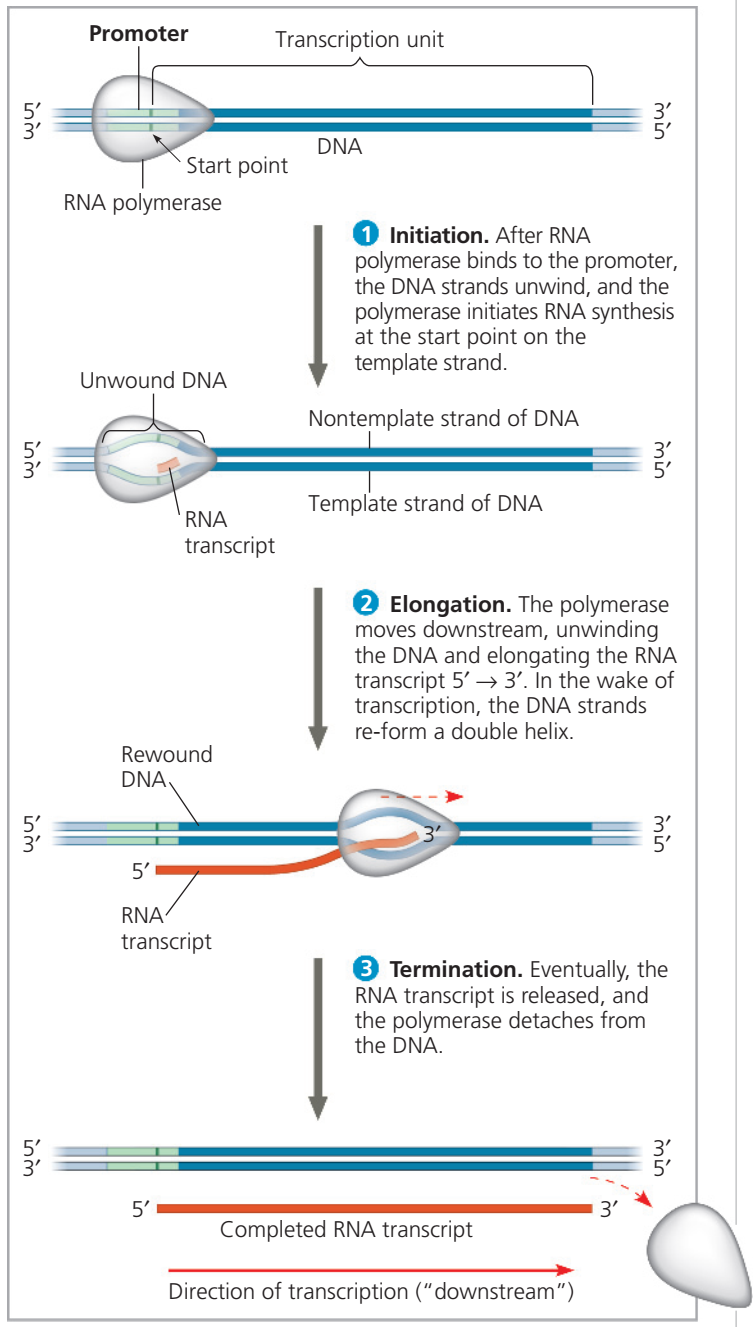
Eşyazımın Moleküler Bileşenleri Bilginin DNAdan hücrenin protein-bireştirici düzeneğine taşınmasını sağlayan iletmen RNA bir genin kalıp telinden eşyazım yoluyla üretilir. RNA polimeraz denilen bir enzim DNAnın iki telini birbirinden ayırır ve DNA kalıp teline tümleyici RNA nükleotidlerini biraraya getirerek RNA polinükleotidi uzatır (Figür 17.8). DNA eşlemlemede işlev gören DNA polimerazlar gibi, RNA polimeraz ancak 3' ucuna ekleme yapabildiği için bir polinükleotidi ancak 5'—3' yönünde birleştirebilir. Ama, DNA polimerazların tersine, RNA polimerazlar bir zinciri sıfırdan başlatabilir; ilk nükleotidi önceden varolan bir primer üzerine ekleme gereksiniminde değildirler.
DNA boyunca özgül nükleotid dizileri bir genin eşyazımının nerede başlayacağını ve nerede sonlanacağını belirtir. RNA polimerazın üzerine bağlanarak eşyazımı başlattığı DNA dizisi başlatıcı (promoter) olarak bilinir; bakteride, eşyazımın sonunu imleyen diziye sonlandırıcı (terminator) denir. (Sonlandırma düzeneği ökaryotlarda başka türlü yer alır). Moleküler biologlar eşyazımın yönüne “aşağıya doğru” (downstream) ve öteki yöne “yukarıya doğru” (upstream) olarak değinirler. Bu terimler DNA ya da RNA içerisindeki nükleotid dizilerinin konumu betimlemek için de kullanılır. Böylece DNAdaki başlatıcı dizinin sonlandırıcıdan yukarıya doğru olduğu söylenir. Başlatıcıdan aşağıya doğru bir RNA molekülüne eşyazımı yapılan DNA parçasına eşyazım birimi denir.
Bakterilerde yalnızca mRNAyı değil ama ayrıca örneğin protein bireşiminde işlev gören ribozomal RNA gibi başka RNA tiplerini de bireştiren tek bir RNA polimeraz tipi vardır. Karşıt olarak, ökaryot çekirdeğinde en az üç tip RNA polimeraz bulunur; ön-mRNA bireşimi için kullanılan birine RNA polimeraz II denir. Öteki RNA polimerazlar proteine çevrilmeyen RNA moleküllerinin eşyazımını yapar. Aşağıdaki tartışmada hem bakterilere hem de ökaryotlara ortak mRNA bireşiminin özellikleri ile başlanacak ve sonra kimi anahtar ayrımları betimlenecektir.
Molecular Components of Transcription Messenger RNA, the carrier of information from DNA to
the cell’s protein-synthesizing machinery, is transcribed
from the template strand of a gene. An enzyme called an
RNA polymerase pries the two strands of DNA apart and
joins together RNA nucleotides complementary to the DNA
template strand, thus elongating the RNA polynucleotide
(Figure 17.8). Like the DNA polymerases that function in
DNA replication, RNA polymerases can assemble a polynucleotide only in its 5' — 3' direction, adding onto its 3'
end. Unlike DNA polymerases, however, RNA polymerases
are able to start a chain from scratch; they don’t need to add
the first nucleotide onto a pre-existing primer.
Specific sequences of nucleotides along the DNA mark
where transcription of a gene begins and ends. The DNA
sequence where RNA polymerase attaches and initiates
transcription is known as the promoter; in bacteria, the
sequence that signals the end of transcription is called the
terminator. (The termination mechanism is different in
eukaryotes; we’ll describe it later.) Molecular biologists refer
to the direction of transcription as “downstream” and the
other direction as “upstream.” These terms are also used to
describe the positions of nucleotide sequences within the
DNA or RNA. Thus, the promoter sequence in DNA is said to
be upstream from the terminator. The stretch of DNA downstream from the promoter that is transcribed into an RNA
molecule is called a transcription unit.
Bacteria have a single type of RNA polymerase that synthesizes not only mRNA but also other types of RNA that function in protein synthesis, such as ribosomal RNA. In contrast,
eukaryotes have at least three types of RNA polymerase in
their nuclei; the one used for pre-mRNA synthesis is called
RNA polymerase II. The other RNA polymerases transcribe
RNA molecules that are not translated into protein. In the
discussion that follows, we start with the features of mRNA
synthesis common to both bacteria and eukaryotes and then
describe some key differences.
Figür 17.8
Eşyazımın evreleri: Başlatma, uzatma ve sonlandırma. Eşyazımın bu genel betimlemesi hem bakteriler hem de ökaryotlar için geçerlidir, ama metinde betimlendiği gibi sonlanmanın ayrıntıları değişiklikler gösterir. Yine, bakteride RNA eşyazımı doğrudan doğruya mRNA olarak kullanılabilirken, bir ökaryotta RNA eşyazımının ilkin işlemden geçmesi gerekir.
Figure 17.8
The stages of transcription: initiation,
elongation, and termination. This general depiction of transcription applies to both bacteria and eukaryotes, but the details of termination differ, as described in the text. Also, in a bacterium, the RNA transcript is immediately usable as mRNA; in a eukaryote, the RNA transcript must first undergo processing.
Bir RNA Eşyazımının Bireşimi Figür 17.8'de gösterildiği ve sonra betimlendiği gibi, eşyazımın üç evresi RNA zincirinin başlatılma, uzatılma ve sonlandırılmasıdır. Evrelere ve betimlenmeleri için kullanılan terimlere alışmak için Figür 17.8'i inceleyin.
Synthesis of an RNA Transcript The three stages of transcription, as shown in Figure 17.8 and described next, are initiation, elongation, and termination of
the RNA chain. Study Figure 17.8 to familiarize yourself with
the stages and the terms used to describe them.
RNA Polimeraz Bağlanması ve Eşyazımın Başlaması
Bir genin başlatıcısı kendi içinde eşyazım başlama noktasını kapsar. Bu nokta RNA polimerazın edimsel olarak mRNA bireşimini başlattığı nükleotiddir. Başlatıcı tipik olarak başlama noktasından yukarıya doğru birçok düzine kadar nükleotid çifti kapsar (Figür 17.9).
Belli proteinler ile etkileşim içinde, RNA polimeraz sağın bir konum ve yönelim ile başlatıcıya bağlanır. Bu kendi payına eşyazımın nerede başlayacağını ve DNA sarmalının iki telinden hangisinin kalıp olarak kullanılacağını belirler.
Bir başlatıcının belli kesimleri RNA polimerazın eşyazımı doğru yerde başlatmasını sağlayacağı bir yolda bağlanması için özellikle önemlidir. Bakterilerde, RNA polimerazın kendisinin bir bölümü bağlatıcıyı özgül olarak tanır ve ona bağlanır. Ökaryotlarda, eşyazım etmenleri denilen belli bir proteinler kümesi RNA polimerazın bağlanmasına ve eşyazımın başlamasına aracılık eder. Ancak eşyazım etmenleri başlatıcıya bağlandıktan sonra RNA polimeraz II ona bağlanır. Başlatıcıya bağlanan bütün bir eşyazım etmenleri ve RNA polimeraz II karmaşasına eşyazım başlatma karmaşası denir. Figür 17.9 bir ökaryotik başlatıcıda başlama karmaşasının oluşumunda eşyazım etmenlerinin ve TATA kutusu denilen önemli bir başlatıcı DNA dizisinin
rolünü gösterir.
Ökaryotik RNA polimeraz II ve eşyazım etmenleri arasındaki etkileşim ökaryotik eşyazımyi denetlemede protein-protein etkileşimlerinin önemini gösteren bir örnektir. Bir kez uygun eşyazım etmenleri başlatıcı DNAya sıkıca eklendikten ve polimeraz DNA üzerinde doğru yönelimde onlara bağlandıktan sonra, enzim başlama noktasında iki DNA telini açar ve kalıbın eşyazımına başlar.
RNA Polymerase Binding and Initiation
of Transcription
The promoter of a gene includes within it the transcription start point—the nucleotide where RNA polymerase actually begins synthesis of the mRNA—and typically extends several dozen or so nucleotide pairs upstream from
the start point (Figure 17.9). Based on interactions with proteins that will be covered shortly, RNA polymerase binds in
a precise location and orientation on the promoter. This in
turn determines where transcription starts and which of the
two strands of the DNA helix is used as the template.
Certain sections of a promoter are especially important
for binding RNA polymerase in a way that ensures that
transcription will begin at the right place. In bacteria, part
of the RNA polymerase itself specifically recognizes and
binds to the promoter. In eukaryotes, a collection of proteins called transcription factors mediate the binding
of RNA polymerase and the initiation of transcription. Only
after transcription factors are attached to the promoter does
RNA polymerase II bind to it. The whole complex of transcription factors and RNA polymerase II bound to the promoter is
called a transcription initiation complex. Figure 17.9
shows the role of transcription factors and a crucial promoter
DNA sequence called the TATA box in forming the initiation
complex at a eukaryotic promoter
The interaction between eukaryotic RNA polymerase II
and transcription factors is an example of the importance
of protein-protein interactions in controlling eukaryotic
transcription. Once the appropriate transcription factors are
firmly attached to the promoter DNA and the polymerase
is bound to them in the correct orientation on the DNA,
the enzyme unwinds the two DNA strands and begins
transcribing the template strand at the start point.
RNA Telinin Uzatılması RNA polimeraz DNA boyunca devinirken, çifte sarmalı çözer ve RNA nükleotidleri ile eşleşmek üzere bir kerede 10 ya da 20 kadar nükleotidi açığa serer (Figür 17.10). Enzim çifte sarmal boyunca ilerlerken büyümekte olan RNA molekülünün 3' ucuna nükleotidler ekler. RNA bireşiminin bu ilerleyen dalgasının peşinde, yeni RNA molekülü DNA kalıbından sıyrılıp uzaklaşır, ve DNA çift sarmalı yeniden oluşur. Eşyazım ökaryotlarda saniyede yaklaşık 40 nükleotid oranında ilerler.
Tek bir gen için birbirini bir konvoydaki taşıtlar gibi izleyen çok sayıda RNA polimeraz molekülü tarafından eşzamanlı olarak eşyazım yapılabilir. Büyümekte olan bir RNA teli her bir polimerazdan sıyrılır ve bu arada her bir yeni telin uzunluğu enzimin kalıp boyunca başlama noktasından ne kadar ilerlediğini gösterir (bkz. Figür 17.23'te mRNA molekülleri). Eşzamanlı olarak tek bir geni eşyazımı yapılmakta olan çok sayıda polimeraz molekülünün kümelenmesi genden eşlenen mRNA miktarını arttırır ve bu büyük miktarlarda kodlanmış molekül üretmede hücreye yardım eder.
Elongation of the RNA Strand As RNA polymerase moves along the DNA, it untwists the
double helix, exposing about 10–20 DNA nucleotides at a
time for pairing with RNA nucleotides (Figure 17.10). The
enzyme adds nucleotides to the 3' end of the growing RNA
molecule as it continues along the double helix. In the wake
of this advancing wave of RNA synthesis, the new RNA molecule peels away from its DNA template, and the DNA double
helix re-forms. Transcription progresses at a rate of about
40 nucleotides per second in eukaryotes.
A single gene can be transcribed simultaneously by several molecules of RNA polymerase following each other like
trucks in a convoy. A growing strand of RNA trails off from
each polymerase, with the length of each new strand reflecting how far along the template the enzyme has traveled from
the start point (see the mRNA molecules in Figure 17.23). The
congregation of many polymerase molecules simultaneously
transcribing a single gene increases the amount of mRNA
transcribed from it, which helps the cell make the encoded
protein in large amounts.
Eşyazımın Sonlandırılması Bakteriler ve ökaryotlar eşyazımı sonlandırma yolunda ayrılır. Bakterilerde, eşyazım DNAdaki bir sonlandırıcı dizi yoluyla ilerler. Eşyazılı sonlandırıcı (bir RNA dizisi) bitiş sinyali olarak işlev görür ve polimerazın DNAdan koparak eşyazımı salmasına neden olur. Eşyazım çeviriden önce daha öte değişki gerektirmez. Ökaryotlarda, RNA polimeraz II DNA üzerinde poliadenilasyon sinyal dizisi denilen bir dizinin eşyazımını yapar. Bu dizi ön-mRNAda bir poliadenilasyon sinyalini (AAUAAA) belirler. Buna bir "sinyal" denir, çünkü bir kez altı RNA nükleotidinden oluşan bu parça görünür görünmez, hemen çekirdekteki beli proteinler tarafından bağlanır. Sonra, AAUAAA'dan 10-35 nükleotid kadar aşağıdaki bir noktada, bu proteinler RNA eşyazımı polimerazdan kopararak ön-mRNAyı salarlar. Sonra ön-mRNA işlemden geçer (sonraki kesimin konusu). O yarılmanın mRNAnın sonunu belirlemesine karşın, RNA polimeraz II eşyazımı sürdürür. Enzimler RNAyı onun yeni açığa serilen 5' ucundan bozundurmaya başlar. Polimeraz enzimler tarafından izlenerek eşyazımı sürdürür ve sonunda enzimler polimeraza yetişince polimeraz DNAdan ayrılır.
Termination of Transcription Bacteria and eukaryotes differ in the way they terminate transcription. In bacteria, transcription proceeds through a terminator sequence in the DNA. The transcribed terminator (an RNA sequence) functions as the termination signal, causing the polymerase to detach from the DNA and release the transcript, which requires no further modification before translation. In eukaryotes, RNA polymerase II transcribes a sequence on the DNA called the polyadenylation signal sequence, which specifies a polyadenylation signal (AAUAAA) in the pre-mRNA. This is called a “signal” because once this stretch of six RNA nucleotides appears, it is immediately bound by certain proteins in the nucleus. Then, at a point about 10–35 nucleotides downstream from the AAUAAA, these proteins cut the RNA transcript free from the polymerase, releasing the pre-mRNA. The pre-mRNA then undergoes processing, the topic of the next section. Although that cleavage marks the end of the mRNA, the RNA polymerase II continues to transcribe. Enzymes begin to degrade the RNA starting at its newly exposed 5' end. The polymerase continues transcribing, pursued by the enzymes, until they catch up to the polymerase and it dissociates from the DNA.
Figür 17.9
Bir ökaryotik başlatıcıda eşyazımın başlaması. Ökaryotik hücrelerde eşyazım etmenleri denilen proteinler RNA polimeraz II ile eşyazımın başlamasına aracılık eder.
Figure 17.9
The initiation of transcription at a eukaryotic
promoter. In eukaryotic cells, proteins called transcription factors
mediate the initiation of transcription by RNA polymerase II.
Figür 17.10
RNA telinin uzatılması
RNA polimeraz DNA kalıp teli boyunca devinerek tümleyici RNA nükleotidlerini büyüyen RNA eşleminin 3' ucuna birleştirir. Polimerazın arkasında, yeni RNA kalıp telden sıyrılır ve bu sonuncusu kalıp-olmayan tel ile yeniden bir çifte sarmal oluşturur.
Figure 17.10
Transcription elongation
RNA polymerase
moves along the DNA template strand, joining complementary RNA
nucleotides to the 3' end of the growing RNA transcript. Behind the
polymerase, the new RNA peels away from the template strand, which
re-forms a double helix with the nontemplate strand.
Figür 17.25
Bir ökaryotik hücredeki eşyazım ve çevirinin bir özeti. Bu çizge bir genden bir polipeptide giden yolu göstermektedir.
DNAdaki her bir gen için yineleyerek birçok özdeş RNA molekülüne eşyazım yapılabilir ve her bir mRNA yineleyerek birçok özdeş polipeptid molekülü vermek üzere çevrilebilir. (Ayrıca, anımsanabilir ki kimi genlerin son ürünleri polipeptidler değil, ama çevrilmeyen tRNA ve rRNA gibi RNA molekülleridir.) Genel olarak, eşyazım ve çeviri adımları bakteriel, arkhaeal ve ökaryotik hücrelerde benzerdir. Başlıca ayrım ökaryotik çekirdekte RNA işleme süreçlerinin bulunmasıdır. Hem eşyazımın hem de çevirinin başlama evrelerinde ve eşyazımın sonlandırılmasında başka önemli ayrımlar bulunur.
Figure 17.25
A summary of transcription and translation in a eukaryotic cell. This diagram shows the path from one gene to
one polypeptide. Each gene in the DNA can be
transcribed repeatedly into many identical RNA
molecules and each mRNA can be translated
repeatedly to yield many identical polypeptide
molecules. (Also, remember that the final
products of some genes are not polypeptides
but RNA molecules that don’t get translated,
including tRNA and rRNA.) In general, the steps
of transcription and translation are similar in
bacterial, archaeal, and eukaryotic cells. The
major difference is the occurrence of RNA
processing in the eukaryotic nucleus. Other
significant differences are found in the initiation
stages of both transcription and translation and
in the termination of transcription.
Eşyazım sürecinin evreleri: Başlatım, uzatım, ve sonlandırım
Şekil eşyazım döngüsünün genel şemasını verir. Gösterilen özellikler hem bakteriel hem de ökaryotik durumlar için geçerlidir.
RNA zincirinin başlangıcını kodlayan DNA nükleotidine eşyazım başlatım sitesi denir ve
“+1” konumu ile belirtilir. Eşyazımın ilerleme yönündeki dizilerinin başlangıç sitesinden aşağıya doğru oldukları söylenir. Benzer olarak, başlama sitesini önceleyen dizilere yukarıya doğru diziler denir. Yukarıya doğru dizilerdeki belirli bir konum belirtilirken, bu negatif bir değer olarak verilir. Aşağıya doğru dizilere pozitif değer yüklenir. (Molecular Biology of the Gene-Pearson, 2013).
🛑 ÖKARYOTİK RNA İŞLEYİMİ 🟡
Ökaryotik RNA işleyimi
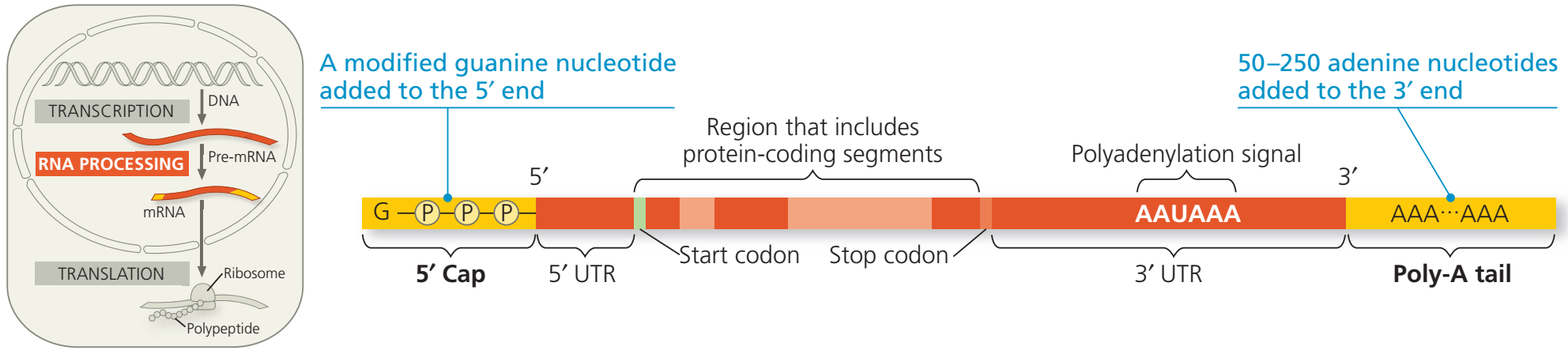
Genetik ileti sitoplazmaya yollanmadan önce, ökaryotik çekirdekte ön-mRNA enzimler tarafından işlenerek değişkiye uğratılır.
İlk olarak, birincil eşyazımın her iki ucu değiştirilir.
İkinci olarak, birçok durumda RNA molekülünün belli iç kesimleri de kesilip atılır ve geriye kalan parçalar biraraya tutturulur (splicing).
Bu değişkiler çeviri için hazır bir mRNA molekülü üretir.
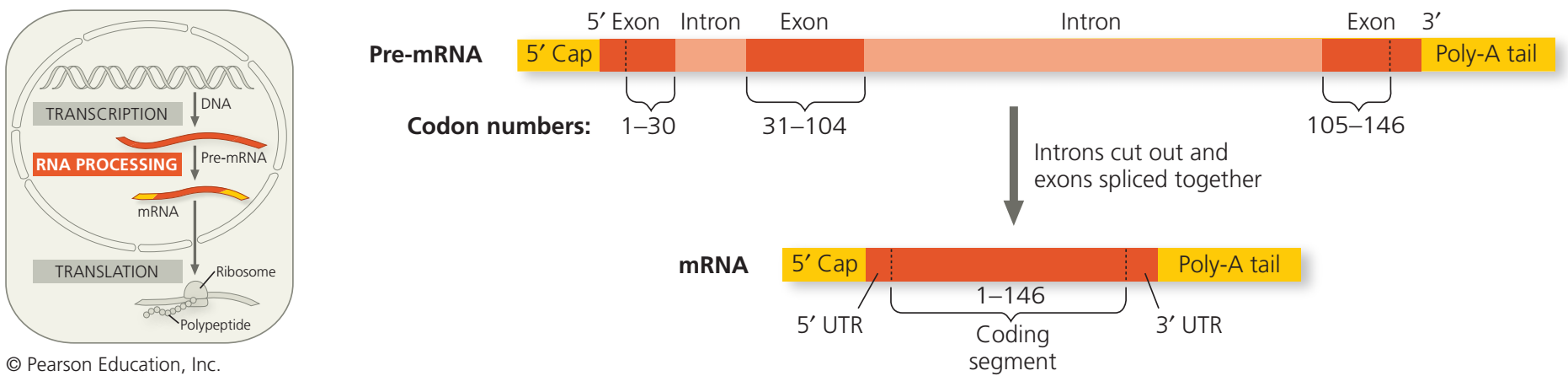
Ökaryotik çekirdekte yer alan RNA işleyiminde ham RNA molekülünde bulunan belli kesimler kesilip atılır.
Bu kes-yapıştır işlemine RNA tutturma (RNA splicing) denir.
Bir insan DNA molekülü boyunca bir eşyazım biriminin ortalama uzunluğu 27.000 nükleotid çifti kadardır (birincil RNA eşyazımının uzunluğu da bu kadardır).
Ortalama 400 amino asit büyüklüğündeki bir protein kodlanmak için RNA molekülünde yalnızca 1.200 nükleotid gerektirir (her bir amino asit bir nükleotidler üçlüsü (kodon) tarafından kodlanır).
Kodonların eşyazımı ve eşlenimi (DNA → RNA → protein).
Buna göre, ökaryotik genler ve bunların RNA eşyazımları kodlama-yapmayan ve çevrilmeyen uzun nükleotid bölgeleri kapsar.
Bu kodlama-yapmaya bölgeler genin ve mRNAnın kodlayıcı bölgeleri arasına dağılmıştır (intronlar (intrude: izinsiz girmek)).
Amino asit dizilerine çevrilerek anlatım bulan kodlayıcı bölgelere exonlar denir (çıkış yapan bölgeler).
(Exonların yalnızca RNAnın uçlarında bulunan UTR bölgeleri proteine çevrilmez.)
RNA polimeraz DNAdan hem exon hem de intronların eşyazımını yapar.
Sitoplazmaya giren mRNA molekülü ön-mRNAnın kısaltılmış bir biçimidir.
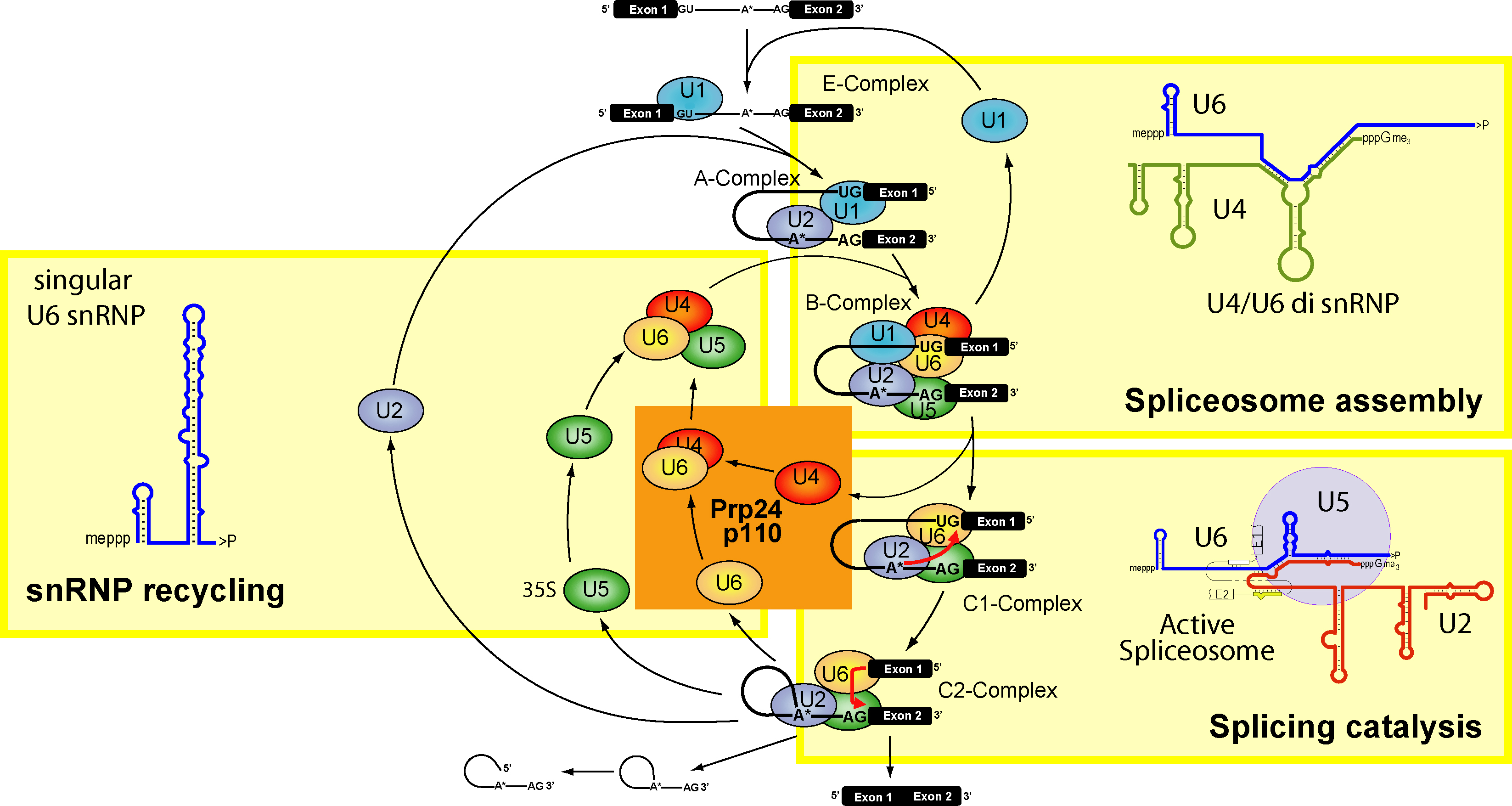
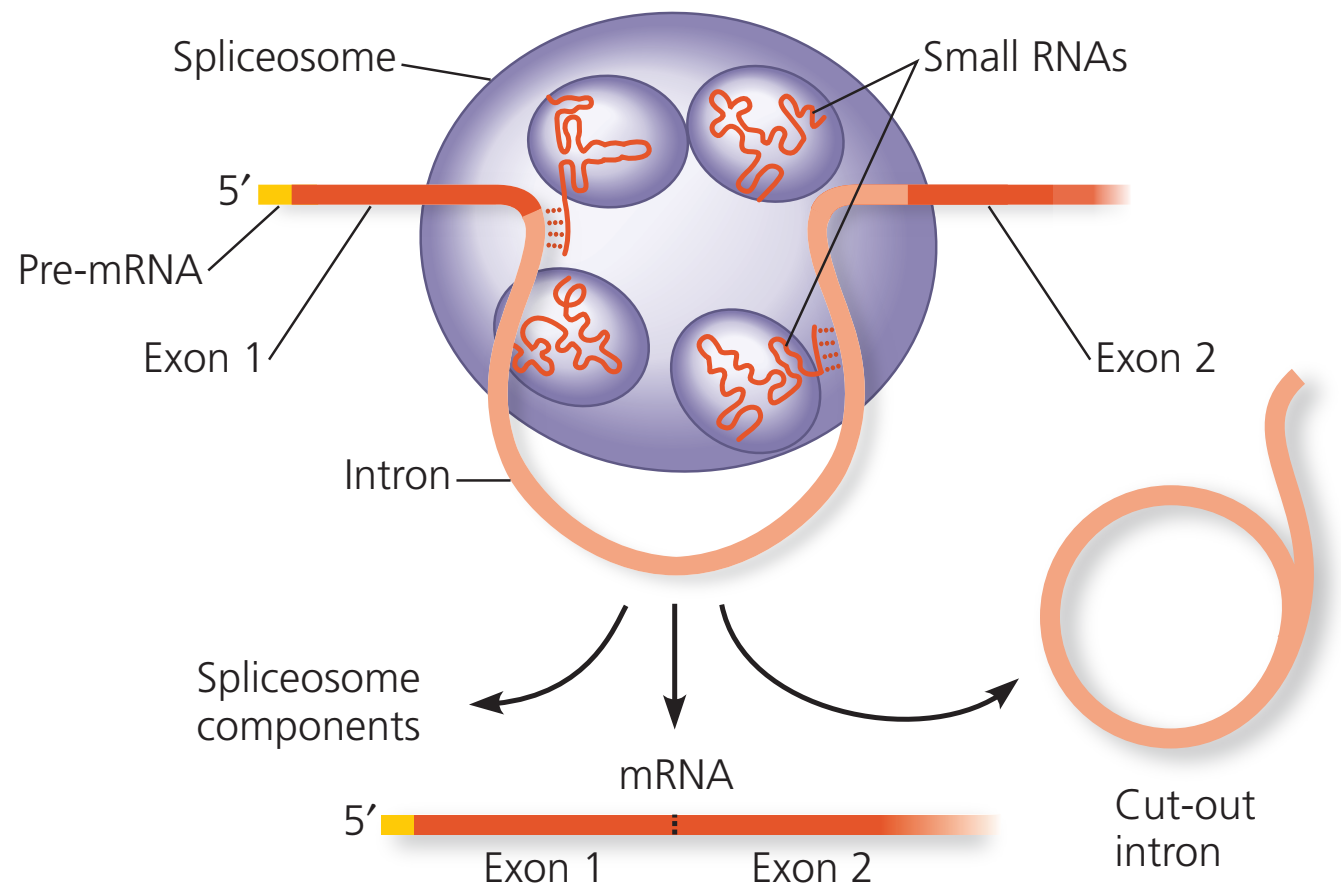
Ön-mRNA tutturma süreci proteinlerden ve küçük RNAlardan oluşan ve splisesom (spliceosome) denilen büyük bir karmaşık yapı tarafından yerine getirilir.
Bu karmaşa bir intron boyunca her bir uçtaki anahtar dizilere aralarında olmak üzere birçok kısa nükleotid dizisine bağlanır.
RNA işleyimi: RNA tutturumu. Burada gösterilen RNA molekülü hemoglobinin polipeptidlerinden biri olan β-globin için kodlama yapar. RNAnın altındaki sayılar kodonları belirtir; β-globin 146 amino asit uzunluğundadır. β-globin geni ve onun ön-mRNA eşyazımı çekirdeği mRNA olarak terk edecek dizilere karşılık düşen üç exon kapsar.(5' UTR ve 3' UTR exonların parçalarıdır çünkü mRNAda kapsanırlar; ama protein için kodlama yapmazlar.) RNA işleyimi sırasında, intronlar kesilip atılır ve exonlar biraraya tutturulur. Birçok gende intronlar exonlardan çok daha uzundur. (“Biology, A Global Approach,” Pearson Education (2020, 343).
Ribonükleoproteinlerin (snRNP) kullanımı ile RNA tutturma süreci.
RNA tutturma işleminde ön-mRNA eşyazımı olgun iletmen RNAya (mRNA) dönüştürülür. Tutturma sırasında, kodlayıcı olmayan intronlar çıkarılır ve kodlayıcı exonlar biraraya tutturulur. Birçok ökaryotik intron durumunda tutturma işlemi splisesom tarafından katalize edilen bir tepkimeler dizisinde yerine getirilir. Splisesom çekirdekte bulunan küçük ribonükleproteinlerin (snRNPler) bir karmaşasıdır. Kendi kendilerini tutturan intronlar, ya da kendilerini ön RNA molekülünden koparma işlemlerinde kendini katalize edebilen ribozimler (katalizör RNA molekülleri) de çekirdekten sitoplazmaya geçer. (W)
🛑 SPLISESOM (SPLICESOME)
SPLISESOM (SPLICESOME)
Splisesom başlıca ökaryotik çekirdekte bulunan büyük ve karmaşık bir moleküler RNA yapısıdır.
Splisesom küçük çekirdek RNAlardan (snRNA) ve yaklaşık olarak 80 proteinden oluşur.
Splisesom eşyazımı yapılan bir ön-mRNA molekülünden intronları ayıklar.
Sürece genel olorak tutturma (splicing) denir. (W)
Splisesom Döngüsü. Tutturma üç ardışık evreye bölünebilir. 1) Splisesomun adım-adım kuruluşu (splisesom toplanımı); 2) yapısal yeniden-düzenleme ve sonraki tutturma tepkimesi ile etkinleştirme (tutturma katalizi); ve 3) tekil alt-birimlerin yeniden kullanıma girişi (snRNP yeniden-döngüsü).
Der Spliceosom-Zyklus. Das Splicing kann in drei aufeinanderfolgende Phasen eingeteilt werden: Der schrittweise Aufbau des Spliceosoms (Spliceosome assembly), die strukturelle Umlagerung und Aktivierung mit der darauffolgenden Splice-Reaktion (Splicing catalysis) und das Recycling der einzelnen Untereinheiten (snRNP recycling)..
Bir ön-mRNA için tutturma işlemini yerine getiren bir splisesom. Çizge bir ön-mRNA eşyazımının bir bölümünü göstermektedir. İntron (pembe) iki exon (kırmızı) tarafından her iki yandan çevrilidir. Splisesom baz çifti içerisindeki küçük RNAlar intron boyunca özgül sitelerde nükleotidler ile baz-çiftleri oluşturur. Sonra, splisesom ön-mRNAnın kesilmesini ve exonların biraraya tutturulmasını katalize eder, intronu hızlı bozunuma bırakır.
🛑 RİBOZİM
RİBOZİM
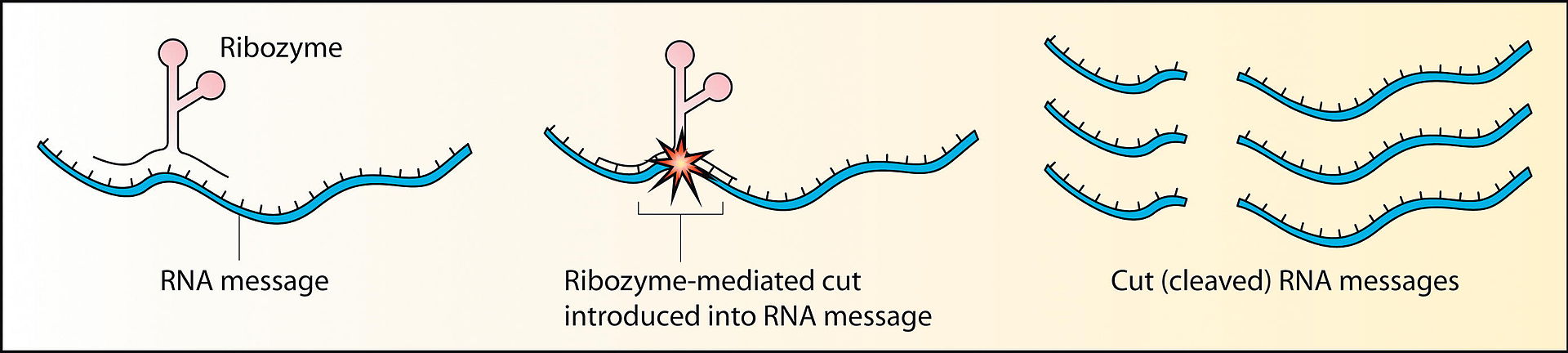
Ribozimler (rbonükleik asit enzimleri) gen anlatımında RNA tutturma gibi özgül biokimyasal tepkimeleri protein enzimlerin eylemine benzer bir yolda katalize edebilen RNA molekülleridir.
RNA hem genetik gereç (DNA gibi), hem de bir kalalisttir (protein enzimler gibi).
Ribozimlerin en yaygın etkinlikleri RNA ve DNA moleküllerini yarma ya da bağlama, ve peptid bağ oluşturmadır.
Ribozom içerisinde, ribozimler büyük ribozomal RNA altbiriminin parçası olarak protein bireşimi sırasında amino asitleri bağlar.
Hücre çekirdeğinde, RNA tutturma gibi çeşitli RNA işleyim tepkimelerine, viral eşlenime ve tRNA bireşimine katılır.
Kimi örgenliklerde RNA tutturması proteinler olmaksızın yer alabilir.
İntron RNA bir ribozim gibi işlev görür ve kendi çıkarılmasını katalize eder.
Silialı protist Tetrahymenada ribozomal RNAnın (rRNA) üretiminde kendini-tutturma yer alır.
Ön-rRNA kendi intronlarını çıkarır.
Ribozomların keşfi tüm biolojik katalistlerin proteinler olduğu düşüncesini çürüttü.
RNA —
a) tek-telli olduğu için, bir RNA molekülünün bir bölgesi aynı moleküldeki bir başka tümleyici bölge ile anti-paralel bir düzen içinde baz-eşleşmesi kurabilir ve bununla enzimatik proteinler gibi üç-boyutlu bir yapı kazanabilir.
b) Enzimatik bir proteindeki belli amino asitler gibi, RNAdaki bazların kimileri katalize katılabilen işlevsel gruplar kapsar.
c) RNAnın başka nükleik asit molekülleri (RNA ya da DNA) ile hidrojen bağı kurma yeteneği katalitik etkinliğine özgüllük katar (örneğin splisesomun RNAsı ve bir birincil RNA eşyazımının RNAsı arasındaki tümleyici baz eşleşmesi ribozimin tutturmayı katalize edeceği bölgeyi tam olarak belirler). (“Campbell Biology,” 2020, s. 344.)
RNAnın ribozim tarafından yarılması.
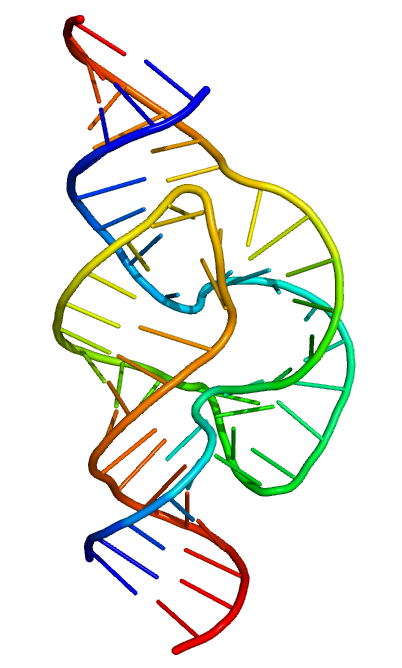
Bir çekiç-başlı ribozimin uzaysal (3D) yapısı. Her bir RNA telinin 5' ucu mavi ve 3' ucu kırmızıdır. Tekil nükleotidler kürdan gibi ve fosfodiester omurga dar bir tüp gibi temsil edilmiştir. (From Protein Data Bank ID 2GOZ.)
🛑 RNA POLİMERAZ
RNA POLİMERAZ
RNA polimeraz ya da ribonükleik asit polimeraz (RNAP) DNAdan RNA bireşimi yapar.
RNA polimeraz bir DNA kalıbından bir RNA polemerinin eşyazım sürecinde katalizörlük eden çok sayıda alt-birimli bir enzimdir.
RNA polimerin nükleotid dizisi kalıp DNAnın nükleotid dizisine tümleyicidir.
DNA kalıbı 3’→ 5′yönünde eşlenir ve RNA polimerin bireşimi 5’→ 3′yönünde yer alır.
Bu RNA teli birincil eşyazım olarak adlandırılır ve hücre içinde işlevsel olabilmek için ilkin işlemden geçmelidir.
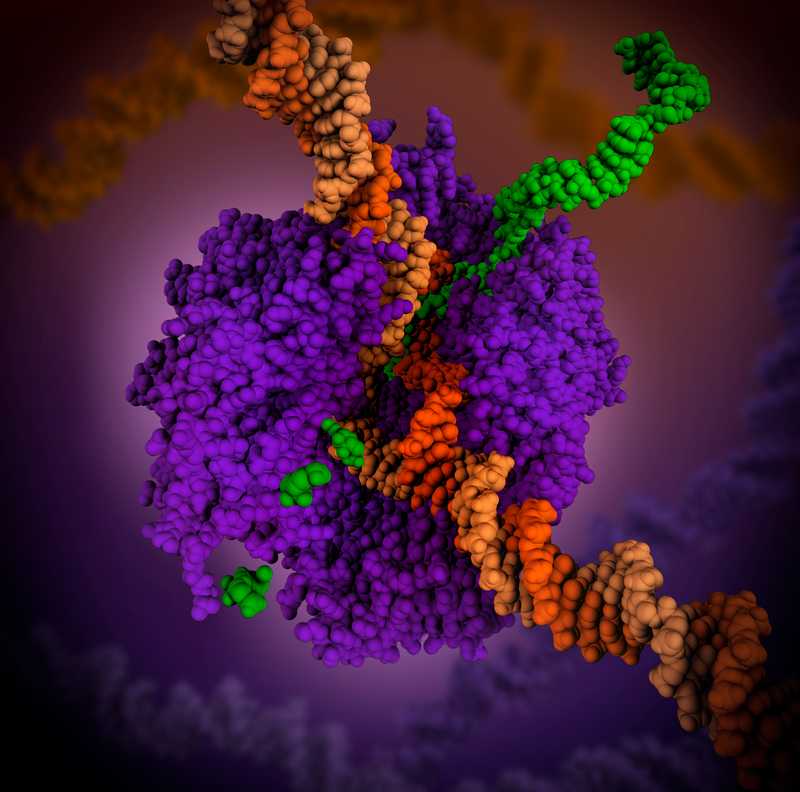
RNA polimeraz (mor) eşyazımın yüreğinde duran bir karmaşık enzimdir. Eşyazım süreci sırasında, enzim DNA çifte sarmalını açar ve tellerden birini (koyu portakal) tek-telli iletmen RNAyı (yeşil) yaratmak için kalıp olarak kullanır. Aynı mRNA molekekülü daha sonra sitoplazmada ribozomlar tarafından protein bireşiminde kullanılır.
RNA polimeraz etkinliğini birçok protein ile etkileşim içinde yerine getirir.
Helikaz enzimini kullanarak, RNAP çift sarmallı DNA molekülünü açar ve DNA tellerinden biri RNA bireşimi (eşyazım) için kalıp olarak kullanılır.
RNA polimerazın DNA açılımını başlatabilmesi için ilkin bir eşyazım etmeni ve onunla birlikte eşyazım aracılık karmaşası DNA bağlanma sitesine bağlanmalıdır.
RNA polimeraz eşyazım sürecinde nükleotidlerin konumlarını denetler, bağlanmalarını ve yeni telin uzatılmasını kolaylaştırır, ve eşyazımın sonlandırılmasında işlevseldir.
RNA polimeraz tarafından üretilen RNAlar işlevsel olarak ya protein kodlayıcı iletmen RNA (mRNA) ya da kodlayıcı-olmayan RNAlardır (bunlara “RNA genleri” denir).
Kimi RNA polimeraz molekülleri dakikada 4.000 baz uzunlukta bir polimerin bireşimini kataliz edebilirler.
Ökaryotlarda, RNA polimeraz 2,4 milyon nükleotid kadar uzun zincirler yapabilir.
RNA genlerinin en az dört işlevsel tipi vardır: —
1) Aktarım RNA (tRNA) — özgül amino asitleri büyüyen poliptid zinrcirine aktarır (çeviri sırasında protein bireşim için ribozomal sitede).
2) Ribozomal RNA (rRNA) — ribozomlara katılır.
3) Mikro RNA (miRNA) — gen etkinliğini düzenler,
4) Katalik RNA (ribozim) — enzimatik olarak etkin RNA molekülü olarak davranır.
Ökaryotik RNA polimerazın her biri ayrı bir RNA altkümesinin bireşimini yapan çok sayıda tipi vardır: —
RNA polimeraz I — bir ön-rRNA 45S bireşimi yapar ve bu olgunlaşarak ribozomun önemli kesimlerini oluşturacaktır.
RNA polimeraz II — mRNAnın öncellerinin, sRNA ve mikroRNAnın bireşimini yapar.
RNA polimeraz III — tRNA, rRNA 5s ve çekirdek ve sitozolde bulunan başka küçük RNAların bireşimini yapar.
RNA polimeraz IV ve V — bitkelerde bulunur ve sRNA yapar.
📹📹📹 TRANSCRIPTION (VİDEO)
📹 04 Transcription / Oxford Academic (Oxford University Press) (VİDEO)
📹 04 Transcription / Oxford Academic (Oxford University Press) (LINK)
In this animation, we explore transcription, the process through which mRNA is synthesised from a DNA template.
NDSU Virtual Cell Animations Project animation 'Transcription'. For more information please see http://vcell.ndsu.edu/animations
📹 Transcription and Translation / Bozeman Science (VİDEO)
📹 Transcription and Translation / Bozeman Science (LINK)
Paul Andersen explains the central dogma of biology. He explains how genes in the DNA are converted to mRNA through the process of transcription. He then explains how ribosomes use this message to convert the mRNA to a functioning protein. He also shows you how to decode a gene by converting the DNA to complementary mRNA and then to the specific amino acids in a protein.
📹 Transcription and Gene Expression / Teachers Pet (VİDEO)
📹 Transcription and Gene Expression / Teachers Pet (LINK)
Learn about the factors effecting gene expression and the control of gene expression during and after transcription in this video!
Various DNA molecular visualizations derived from x-ray crystallography and other data sets, and imbued with dynamic movement that suggest brownian motion.
These molecular animations were created for a major trans-national production effort to raise awareness, educate and promote DNA science to the wider community, coinciding with the 50th anniversary of the discovery of the double helix.
📹 Transcription vs. Translation / Beverly Biology (VİDEO)
📹 Transcription vs. Translation / Beverly Biology (LINK)
Learn the basic concepts behind transcription and translation in this quick video.
📹 RNA and Transcription (updated) / Beverly Biology (VİDEO)
📹 RNA and Transcription (updated) / Beverly Biology (LINK)
📹 Promoters and Transcription (2016) / Alex Lee (VİDEO)
📹 Promoters and Transcription (2016) / Alex Lee (LINK)
7.2 Transcription and gene expression:Promoters and Transcription
Understanding that:
- Transcription require promoters; it is a region of DNA that initiates transcription of a particular gene.
- Promoters are located near the transcription start sites of genes
- Promoters are an example of non-coding DNA with a function
Before being used in translation, mRNA must be spliced. During splicing, introns are removed and the translatable exons that remain are spliced into a single strand of mRNA.
📹 RNA splicing and Spliceosome / Frank Lectures (VİDEO)
📹 RNA splicing and Spliceosome / Frank Lectures (LINK)
RNA splicing
Spliceosome
📹📹📹 POST-TRANSCRIPTIONAL (VİDEO)
📹 Post-transcriptional regulation / Khan Academy (VİDEO)
📹 Post-transcriptional regulation / Khan Academy (LINK)
Transcription, the synthesis of RNA from DNA. Genetic information flows from DNA into protein, the substance that gives an organism its form. This flow of information occurs through the sequential processes of transcription (DNA to RNA) and translation (RNA to protein). Transcription occurs when there is a need for a particular gene product at a specific time or in a specific tissue.
Genes are made up of promoter regions and alternating regions of introns (noncoding sequences) and exons (coding sequences). The production of a functional protein involves the transcription of the gene from DNA into RNA, the removal of introns and splicing together of exons, the translation of the spliced RNA sequences into a chain of amino acids, and the posttranslational modification of the protein molecule.
During transcription, only one strand of DNA is usually copied. This is called the template strand, and the RNA molecules produced are single-strandedmessenger RNAs (mRNAs). The DNA strand that would correspond to the mRNA is called the coding or sense strand. In eukaryotes (organisms that possess a nucleus) the initial product of transcription is called a pre-mRNA. Pre-mRNA is extensively edited through splicing before the mature mRNA is produced and ready for translation by the ribosome, the cellular organelle that serves as the site of protein synthesis. Transcription of any one gene takes place at the chromosomal location of that gene, which is a relatively short segment of the chromosome. The active transcription of a gene depends on the need for the activity of that particular gene in a specific cell or tissue or at a given time.
Small segments of DNA are transcribed into RNA by the enzymeRNA polymerase, which achieves this copying in a strictly controlled process. The first step is to recognize a specific sequence on DNA called a promoter that signifies the start of the gene. The two strands of DNA become separated at this point, and RNA polymerase begins copying from a specific point on one strand of the DNA using a special type of sugar-containing nucleoside called ribonucleoside 5’-triphosphate to begin the growing chain. Additional ribonucleoside triphosphates are used as the substrate, and, by cleavage of their high-energy phosphate bond, ribonucleoside monophosphates are incorporated into the growing RNA chain. Each successive ribonucleotide is directed by the complementary base pairing rules of DNA. For example, a C (cytosine) in DNA directs the incorporation of a G (guanine) into RNA. Likewise, a G in DNA is copied into a C in RNA, a T (thymine) into an A (adenine), and an A into a U (uracil; RNA contains U in place of the T of DNA). Synthesis continues until a termination signal is reached, at which point the RNA polymerase drops off the DNA, and the RNA molecule is released.
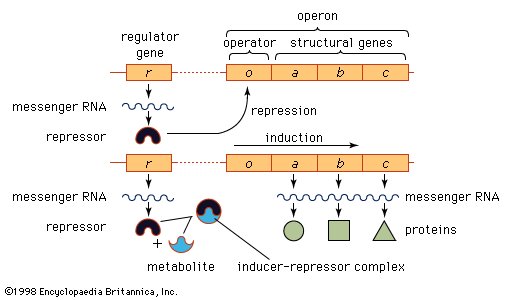
Ahead of many genes in prokaryotes (organisms that lack a nucleus), there are signals called “operators” (seeoperons) where specialized proteins called repressors bind to the DNA just upstream of the start point of transcription and prevent access to the DNA by RNA polymerase. These repressor proteins thus prevent transcription of the gene by physically blocking the action of the RNA polymerase. Typically, repressors are released from their blocking action when they receive signals from other molecules in the cell indicating that the gene needs to be expressed. Ahead of some prokaryotic genes are signals to which activator proteins bind to stimulate transcription.
Model of the operon and its relation to the regulator gene.
Transcription in eukaryotes is more complicated than in prokaryotes. First, the RNA polymerase of higher organisms is a more complicated enzyme than the relatively simple five-subunit enzyme of prokaryotes. In addition, there are many more accessory factors that help to control the efficiency of the individual promoters. These accessory proteins are called transcription factors and typically respond to signals from within the cell that indicate whether transcription is required. In many human genes, several transcription factors may be needed before transcription can proceed efficiently. A transcription factor can cause either repression or activation of gene expression in eukaryotes.
Simplified diagram of mRNA synthesis and processing. Enzymes not shown.
🔎
RNA polymerase generates a transcription bubble, which separates the two strands of the DNA helix. This is done by breaking the hydrogen bonds between complementary DNA nucleotides.
RNA polymerase adds RNA nucleotides (which are complementary to the nucleotides of one DNA strand).
RNA sugar-phosphate backbone forms with assistance from RNA polymerase to form an RNA strand.
Hydrogen bonds of the RNA–DNA helix break, freeing the newly synthesized RNA strand.
The RNA may remain in the nucleus or exit to the cytoplasm through the nuclear pore complex.
The stretch of DNA transcribed into an RNA molecule is called a transcription unit and encodes at least one gene. If the gene encodes a protein, the transcription produces messenger RNA (mRNA); the mRNA, in turn, serves as a template for the protein's synthesis through translation. Alternatively, the transcribed gene may encode for non-coding RNA such as microRNA,ribosomal RNA (rRNA), transfer RNA (tRNA), or enzymatic RNA molecules called ribozymes. Overall, RNA helps synthesize, regulate, and process proteins; it therefore plays a fundamental role in performing functions within a cell.
In virology, the term may also be used when referring to mRNA synthesis from an RNA molecule (i.e., RNA replication). For instance, the genome of a negative-sense single-stranded RNA (ssRNA -) virus may be template for a positive-sense single-stranded RNA (ssRNA +). This is because the positive-sense strand contains the information needed to translate the viral proteins for viral replication afterwards. This process is catalyzed by a viral RNA replicase.
A DNA transcription unit encoding for a protein may contain both a coding sequence, which will be translated into the protein, and regulatory sequences, which direct and regulate the synthesis of that protein. The regulatory sequence before ("upstream" from) the coding sequence is called the five prime untranslated region (5'UTR); the sequence after ("downstream" from) the coding sequence is called the three prime untranslated region (3'UTR).
As opposed to DNA replication, transcription results in an RNA complement that includes the nucleotide uracil (U) in all instances where thymine (T) would have occurred in a DNA complement.
Only one of the two DNA strands serve as a template for transcription. The antisense strand of DNA is read by RNA polymerase from the 3' end to the 5' end during transcription (3' → 5'). The complementary RNA is created in the opposite direction, in the 5' → 3' direction, matching the sequence of the sense strand with the exception of switching uracil for thymine. This directionality is because RNA polymerase can only add nucleotides to the 3' end of the growing mRNA chain. This use of only the 3' → 5' DNA strand eliminates the need for the Okazaki fragments that are seen in DNA replication. This also removes the need for an RNA primer to initiate RNA synthesis, as is the case in DNA replication.
The non-template (sense) strand of DNA is called the coding strand, because its sequence is the same as the newly created RNA transcript (except for the substitution of uracil for thymine). This is the strand that is used by convention when presenting a DNA sequence.
Transcription has some proofreading mechanisms, but they are fewer and less effective than the controls for copying DNA. As a result, transcription has a lower copying fidelity than DNA replication.
Transcription begins with the binding of RNA polymerase, together with one or more general transcription factors, to a specific DNA sequence referred to as a "promoter" to form an RNA polymerase-promoter "closed complex". In the "closed complex" the promoter DNA is still fully double-stranded.
RNA polymerase, assisted by one or more general transcription factors, then unwinds approximately 14 base pairs of DNA to form an RNA polymerase-promoter "open complex". In the "open complex" the promoter DNA is partly unwound and single-stranded. The exposed, single-stranded DNA is referred to as the "transcription bubble."
RNA polymerase, assisted by one or more general transcription factors, then selects a transcription start site in the transcription bubble, binds to an initiating NTP and an extending NTP (or a short RNA primer and an extending NTP) complementary to the transcription start site sequence, and catalyzes bond formation to yield an initial RNA product.
In bacteria, RNA polymerase holoenzyme consists of five subunits: 2 α subunits, 1 β subunit, 1 β' subunit, and 1 ω subunit. In bacteria, there is one general RNA transcription factor known as a sigma factor. RNA polymerase core enzyme binds to the bacterial general transcription (sigma) factor to form RNA polymerase holoenzyme and then binds to a promoter. (RNA polymerase is called a holoenzyme when sigma subunit is attached to the core enzyme which is consist of 2 α subunits, 1 β subunit, 1 β' subunit only).
In archaea and eukaryotes, RNA polymerase contains subunits homologous to each of the five RNA polymerase subunits in bacteria and also contains additional subunits. In archaea and eukaryotes, the functions of the bacterial general transcription factor sigma are performed by multiple general transcription factors that work together. In archaea, there are three general transcription factors: TBP,TFB, and TFE. In eukaryotes, in RNA polymerase II-dependent transcription, there are six general transcription factors: TFIIA,TFIIB (an ortholog of archaeal TFB), TFIID (a multisubunit factor in which the key subunit, TBP, is an ortholog of archaeal TBP), TFIIE (an ortholog of archaeal TFE), TFIIF, and TFIIH. The TFIID is the first component to bind to DNA due to binding of TBP, while TFIIH is the last component to be recruited. In archaea and eukaryotes, the RNA polymerase-promoter closed complex is usually referred to as the "preinitiation complex."
Transcription initiation is regulated by additional proteins, known as activators and repressors, and, in some cases, associated coactivators or corepressors, which modulate formation and function of the transcription initiation complex.
After the first bond is synthesized, the RNA polymerase must escape the promoter. During this time there is a tendency to release the RNA transcript and produce truncated transcripts. This is called abortive initiation, and is common for both eukaryotes and prokaryotes. Abortive initiation continues to occur until an RNA product of a threshold length of approximately 10 nucleotides is synthesized, at which point promoter escape occurs and a transcription elongation complex is formed.
Mechanistically, promoter escape occurs through DNA scrunching, providing the energy needed to break interactions between RNA polymerase holoenzyme and the promoter.
In bacteria, it was historically thought that the sigma factor is definitely released after promoter clearance occurs. This theory had been known as the obligate release model however later data showed that upon and following promoter clearance, the sigma factor is released according to a stochastic model known as the stochastic release model.
In eukaryotes, at an RNA polymerase II-dependent promoter, upon promoter clearance, TFIIH phosphorylates serine 5 on the carboxy terminal domain of RNA polymerase II, leading to the recruitment of capping enzyme (CE). The exact mechanism of how CE induces promoter clearance in eukaryotes is not yet known.
One strand of the DNA, the template strand (or noncoding strand), is used as a template for RNA synthesis. As transcription proceeds, RNA polymerase traverses the template strand and uses base pairing complementarity with the DNA template to create an RNA copy (which elongates during the traversal). Although RNA polymerase traverses the template strand from 3' → 5', the coding (non-template) strand and newly formed RNA can also be used as reference points, so transcription can be described as occurring 5' → 3'. This produces an RNA molecule from 5' → 3', an exact copy of the coding strand (except that thymines are replaced with uracils, and the nucleotides are composed of a ribose (5-carbon) sugar where DNA has deoxyribose (one fewer oxygen atom) in its sugar-phosphate backbone).
Simple diagram of transcription elongation.
mRNA transcription can involve multiple RNA polymerases on a single DNA template and multiple rounds of transcription (amplification of particular mRNA), so many mRNA molecules can be rapidly produced from a single copy of a gene. The characteristic elongation rates in prokaryotes and eukaryotes are about 10-100 nts/sec. In eukaryotes, however, nucleosomes act as major barriers to transcribing polymerases during transcription elongation. In these organisms, the pausing induced by nucleosomes can be regulated by transcription elongation factors such as TFIIS.
Elongation also involves a proofreading mechanism that can replace incorrectly incorporated bases. In eukaryotes, this may correspond with short pauses during transcription that allow appropriate RNA editing factors to bind. These pauses may be intrinsic to the RNA polymerase or due to chromatin structure.
Bacteria use two different strategies for transcription termination – Rho-independent termination and Rho-dependent termination. In Rho-independent transcription termination, RNA transcription stops when the newly synthesized RNA molecule forms a G-C-rich hairpin loop followed by a run of Us. When the hairpin forms, the mechanical stress breaks the weak rU-dA bonds, now filling the DNA–RNA hybrid. This pulls the poly-U transcript out of the active site of the RNA polymerase, terminating transcription. In the "Rho-dependent" type of termination, a protein factor called "Rho" destabilizes the interaction between the template and the mRNA, thus releasing the newly synthesized mRNA from the elongation complex.
Transcription termination in eukaryotes is less well understood than in bacteria, but involves cleavage of the new transcript followed by template-independent addition of adenines at its new 3' end, in a process called polyadenylation.
In vertebrates, the majority of gene promoters contain a CpG island with numerous CpG sites. When many of a gene's promoter CpG sites are methylated the gene becomes inhibited (silenced). Colorectal cancers typically have 3 to 6 driver mutations and 33 to 66 hitchhiker or passenger mutations. However, transcriptional inhibition (silencing) may be of more importance than mutation in causing progression to cancer. For example, in colorectal cancers about 600 to 800 genes are transcriptionally inhibited by CpG island methylation (see regulation of transcription in cancer). Transcriptional repression in cancer can also occur by other epigenetic mechanisms, such as altered expression of microRNAs. In breast cancer, transcriptional repression of BRCA1 may occur more frequently by over-expressed microRNA-182 than by hypermethylation of the BRCA1 promoter (see Low expression of BRCA1 in breast and ovarian cancers).
Active transcription units are clustered in the nucleus, in discrete sites called transcription factories or euchromatin. Such sites can be visualized by allowing engaged polymerases to extend their transcripts in tagged precursors (Br-UTP or Br-U) and immuno-labeling the tagged nascent RNA. Transcription factories can also be localized using fluorescence in situ hybridization or marked by antibodies directed against polymerases. There are ~10,000 factories in the nucleoplasm of a HeLa cell, among which are ~8,000 polymerase II factories and ~2,000 polymerase III factories. Each polymerase II factory contains ~8 polymerases. As most active transcription units are associated with only one polymerase, each factory usually contains ~8 different transcription units. These units might be associated through promoters and/or enhancers, with loops forming a "cloud" around the factor.
A molecule that allows the genetic material to be realized as a protein was first hypothesized by François Jacob and Jacques Monod.Severo Ochoa won a Nobel Prize in Physiology or Medicine in 1959 for developing a process for synthesizing RNA in vitro with polynucleotide phosphorylase, which was useful for cracking the genetic code. RNA synthesis by RNA polymerase was established in vitro by several laboratories by 1965; however, the RNA synthesized by these enzymes had properties that suggested the existence of an additional factor needed to terminate transcription correctly.
In 1972, Walter Fiers became the first person to actually prove the existence of the terminating enzyme.
MS2 tagging: by incorporating RNA stem loops, such as MS2, into a gene, these become incorporated into newly synthesized RNA. The stem loops can then be detected using a fusion of GFP and the MS2 coat protein, which has a high affinity, sequence-specific interaction with the MS2 stem loops. The recruitment of GFP to the site of transcription is visualized as a single fluorescent spot. This new approach has revealed that transcription occurs in discontinuous bursts, or pulses (see Transcriptional bursting). With the notable exception of in situ techniques, most other methods provide cell population averages, and are not capable of detecting this fundamental property of genes.
Northern blot: the traditional method, and until the advent of RNA-Seq, the most quantitative
RNA-Seq: applies next-generation sequencing techniques to sequence whole transcriptomes, which allows the measurement of relative abundance of RNA, as well as the detection of additional variations such as fusion genes, post-transcriptional edits and novel splice sites
Single cell RNA-Seq: amplifies and reads partial transcriptomes from isolated cells, allowing for detailed analyses of RNA in tissues, embryos, and cancers
Electron micrograph of transcription of ribosomal RNA. The forming ribosomal RNA strands are visible as branches from the main DNA strand.
Some viruses (such as HIV, the cause of AIDS), have the ability to transcribe RNA into DNA. HIV has an RNA genome that is reverse transcribed into DNA. The resulting DNA can be merged with the DNA genome of the host cell. The main enzyme responsible for synthesis of DNA from an RNA template is called reverse transcriptase.
Electron micrograph of transcription of ribosomal RNA. The forming ribosomal RNA strands are visible as branches from the main DNA strand.
In the case of HIV, reverse transcriptase is responsible for synthesizing a complementary DNA strand (cDNA) to the viral RNA genome. The enzyme ribonuclease H then digests the RNA strand, and reverse transcriptase synthesises a complementary strand of DNA to form a double helix DNA structure ("cDNA"). The cDNA is integrated into the host cell's genome by the enzyme integrase, which causes the host cell to generate viral proteins that reassemble into new viral particles. In HIV, subsequent to this, the host cell undergoes programmed cell death, or apoptosis of T cells. However, in other retroviruses, the host cell remains intact as the virus buds out of the cell.
Some eukaryotic cells contain an enzyme with reverse transcription activity called telomerase. Telomerase is a reverse transcriptase that lengthens the ends of linear chromosomes. Telomerase carries an RNA template from which it synthesizes a repeating sequence of DNA, or "junk" DNA. This repeated sequence of DNA is called a telomere and can be thought of as a "cap" for a chromosome. It is important because every time a linear chromosome is duplicated, it is shortened. With this "junk" DNA or "cap" at the ends of chromosomes, the shortening eliminates some of the non-essential, repeated sequence rather than the protein-encoding DNA sequence, that is farther away from the chromosome end.
Telomerase is often activated in cancer cells to enable cancer cells to duplicate their genomes indefinitely without losing important protein-coding DNA sequence. Activation of telomerase could be part of the process that allows cancer cells to become immortal. The immortalizing factor of cancer via telomere lengthening due to telomerase has been proven to occur in 90% of all carcinogenic tumors in vivo with the remaining 10% using an alternative telomere maintenance route called ALT or Alternative Lengthening of Telomeres.
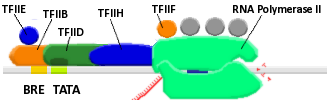
Transcription factors. In the middle part above the promoter, the pink color part of the transcription factors are the General Transcription Factors.
🔎
General transcription factors (GTFs), also known as basal transcriptional factors, are a class of proteintranscription factors that bind to specific sites (promoter) on DNA to activate transcription of genetic information from DNA to messenger RNA. GTFs, RNA polymerase, and the mediator (a multi-protein complex) constitute the basic transcriptional apparatus that first bind to the promoter, then start transcription. GTFs are also intimately involved in the process of gene regulation, and most are required for life.
A transcription factor is a protein that binds to specific DNA sequences (enhancer or promoter), either alone or with other proteins in a complex, to control the rate of transcription of genetic information from DNA to messenger RNA by promoting (serving as an activator) or blocking (serving as a repressor) the recruitment of RNA polymerase. As a class of protein, general transcription factors bind to promoters along the DNA sequence or form a large transcription preinitiation complex to activate transcription. General transcription factors are necessary for transcription to occur.
In bacteria, transcription initiation requires an RNA polymerase and a single GTF: sigma factor.
In archaea and eukaryotes, transcription initiation requires an RNA polymerase and a set of multiple GTFs to form a transcription preinitiation complex. The Transcription initiation by eukaryotic RNA polymerase II involves the following GTFs:
A sigma factor is a protein needed only for initiation of RNA synthesis in bacteria. Sigma factors provide promoter recognition specificity to the RNA polymerase (RNAP) and contribute to DNA strand separation, then dissociating from the RNA polymerase core enzyme following transcription initiation. The RNA polymerase core associates with the sigma factor to form RNA polymerase holoenzyme. Sigma factor reduces the affinity of RNA polymerase for nonspecific DNA while increasing specificity for promoters, allowing transcription to initiate at correct sites. The core enzyme of RNA polymerase has five subunits (protein subunits) (~400 kDa). Because of the RNA polymerase association with sigma factor, the complete RNA polymerase therefore has 6 subunits: the sigma subunit-in addition to the two alpha (α), one beta (β), one beta prime (β'), and one omega (ω) subunits that make up the core enzyme(~450 kDa). In addition, many bacteria can have multiple alternative σ factors. The level and activity of the alternative σ factors are highly regulated and can vary depending on environmental or developmental signals.
The transcription preinitiation complex is a large complex of proteins that is necessary for the transcription of protein-coding genes in eukaryotes and archaea. It attaches to the promoter of the DNA (e.i., TATA box) and helps position the RNA polymerase II to the gene transcription start sites, denatures the DNA, and then starts transcription.
The assembly of transcription preinitiation complex follows these steps:
TATA binding protein (TBP), a subunit of TFIID (the largest GTF) binds to the promoter (TATA box), creating a sharp bend in the promoter DNA. Then the TBP-TFIIA interactions recruit TFIIA to the promoter.
TBP-TFIIB interactions recruit TFIIB to the promoter. RNA polymerase II and TFIIF assemble to form the Polymerase II complex. TFIIB helps the Pol II complex bind correctly.
TFIIE and TFIIH then bind to the complex and form the transcription preinitiation complex. TFIIA/B/E/H leave once RNA elongation begins. TFIID will stay until elongation is finished.
Subunits within TFIIH that have ATPase and helicase activity create negative superhelical tension in the DNA. This negative superhelical tension causes approximately one turn of DNA to unwind and form the transcription bubble.
The template strand of the transcription bubble engages with the RNA polymerase II active site, then RNA synthesis starts.


.png)
{kind=link}